
SODIUM BICARBONATE ON GROWTH PERFORMANCE, BLOOD BIOCHEMICAL PARAMERERS, AND INTESTINAL DAMAGE OF LARGEMOUTH BASS (MICROPTERUS SALMOIDES) UNDER CHRONIC HEAT STRESS
-
摘要:
为了探究碳酸氢钠(NaHCO3)对慢性高温胁迫下大口黑鲈(Micropterus salmoides)肠道损伤的影响, 实验以大口黑鲈幼鱼(20.26±0.08) g为研究对象, 分别投喂含有或不含碳酸氢钠(NaHCO3, 5 g/kg)的饲料8周。结果表明, 在慢性高温胁迫下, NaHCO3组特定生长率与对照组相比无显著差异, 但饲料效率、蛋白质沉积率和蛋白质效率, 以及肠道淀粉酶和胰蛋白酶活性显著提高。NaHCO3的添加显著提高了肠道总抗氧化能力(T-AOC)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GPx)活性, 并上调了cat和gpx1a基因的表达, 下调了keap1的基因表达。同时, NaHCO3显著下调了促炎症细胞因子(il-1β、tnfα、il15)的表达, 上调了抗炎细胞因子il10的表达。此外, NaHCO3组大口黑鲈肠道的hsp90、hsp70和hsp60基因的表达均显著低于对照组。肠道组织学分析表明, 高温胁迫引起了大口黑鲈肠道损伤, 饲料中添加NaHCO3显著改善了肠道组织形态, 并上调了肠道zo1、jam、occludin、muc2的表达。综上所述, 饲料中添加5 g/kg NaHCO3通过提高肠道抗氧化能力、改善肠道炎症和肠道屏障功能来有效缓解慢性高温胁迫诱导的大口黑鲈肠道损伤。研究为改善鱼类在高温胁迫下的肠道健康提供了新的视角。
Abstract:In order to investigate the effect of sodium bicarbonate (NaHCO3) on intestinal damage in largemouth bass (Micropterus salmoides) under chronic high-temperature stress, juvenile largemouth bass (20.26±0.08) g were fed diets with or without sodium bicarbonate (NaHCO3, 5 g/kg) for 8 weeks. The results showed that there was no significant difference in the specific growth rate in the NaHCO3 group compared to the control group at chronic high temperatures. However, the NaHCO3 group exhibited significantly improved feed efficiency, protein deposition rate, and protein efficiency, along with a marked increase in intestinal amylase and trypsin activities. The addition of NaHCO3 also significantly enhanced the intestinal total antioxidant capacity (T-AOC), catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx) activities. Furthermore, NaHCO3 up-regulated the expression of cat and gpx1a genes and down-regulated the gene expression of keap1. Meanwhile, the expressions of pro-inflammatory cytokines (il-1β, tnfα, il15) were significantly down-regulated in the group, while the expression of anti-inflammatory cytokine il10 was up-regulated. In addition, the expression of hsp90, hsp70, and hsp60 genes in the intestines of largemouth bass in the NaHCO3 group was significantly lower than that in the control group. Histologic analysis of the intestinal tract showed that high-temperature stress induced intestinal damage in largemouth bass, and the addition of NaHCO3 significantly improved the intestinal tissue morphology and up-regulated the expression of intestinal zo1, jam, occludin, and muc2. In conclusion, the addition of 5 g/kg NaHCO3 to the feed effectively alleviated chronic high-temperature stress-induced intestinal damage in largemouth bass by improving intestinal antioxidant capacity, intestinal inflammation, and intestinal barrier function. These findings may provide a new perspective to improve the intestinal health of fish under high-temperature stress.
-
Keywords:
- NaHCO3 /
- Heat stress /
- Growth performance /
- Gut health /
- Largemouth bass
-
在水产养殖过程中, 养殖水体温度被认为是最关键的环境因素之一。随着全球气候变暖的加剧, 养殖水温逐渐升高, 鱼类的热应激程度也随之增加[1]。近年来, 中国多个省份在夏季6—8月期间气温超过40℃, 地面温度甚至超过45℃[2]。持续的极端高温使得水温超过了鱼类的耐热限度, 影响其生长与健康[3]。以华南地区为例, 夏季室外池塘养殖水温常常升高至35℃或更高, 导致水产养殖动物摄食量显著下降, 生长速度减缓, 抗病能力降低, 进而严重影响经济效益[4, 5]。
大口黑鲈(Micropterus salmoides), 鲈形目棘臀鱼科黑鲈属, 1983年由北美引进, 因其肉质鲜嫩、营养丰富、生长迅速且易于捕捞, 具有较高的经济价值, 是优良的淡水养殖品种之一[6]。在高温季节, 特别是水温超过28℃时, 大口黑鲈容易出现热应激反应, 免疫力下降, 易感染疾病。同时, 高温导致溶氧不足和水质恶化问题进一步加剧, 进而引发病害暴发和经济损失[7]。研究表明, 在高温环境下, 鱼类的肠道结构容易受到破坏, 进而导致肠道菌群失衡, 引发严重的生理功能障碍[8]。同时高温胁迫会损害大口黑鲈的肠道健康, 导致其营养吸收能力下降[9]。慢性高温应激是诱发鱼类肠炎的主要因素之一, 高温使得大口黑鲈小肠上皮细胞的完整性易受损, 导致肠道黏膜通透性增加, 致病菌容易侵入, 引发炎症反应[10, 11]。
营养调控是指通过调整饲料成分等促进动物营养吸收、增强免疫力等过程[12]。碳酸氢钠(NaHCO3)作为一种无机盐, 具有调节促生长、抗氧化、促进消化、增强免疫力和缓解热应激等多重功能[13]。研究表明, NaHCO3通过改善水质口感和促进饮水量, 有效提升热应激条件下公鸡(Gallus)的生长性能和饲料利用效率[14]。同时, 在肉鸡(Gallus domesticus)中添加NaHCO3显著提高了日增重, 并缓解了慢性高温胁迫引起的呼吸性碱中毒[15]。此外, 作为一种弱碱, NaHCO3可提高机体血液pH, 进而中和代谢物中的酸性物质[16]。NaHCO3显著改善了羔羊(Agnus dei)和羊(Capra hircus L.)的生长性能, 并有效降低了瘤胃pH[17, 18]。在山羊中联合使用铬(1.5 mg/d)和NaHCO3(15 g/kg)显著提高了生长性能, 并通过缓解热胁迫的负面影响, 改善了饲料利用率[19]。尽管NaHCO3在多种动物中的应用已有一定研究, 但在鱼类中的相关研究仍较为有限。本文推测NaHCO3可能提高大口黑鲈耐高温能力。为了验证我们的假设, 我们探讨了在慢性高温胁迫下, 添加5 g/kg NaHCO3对大口黑鲈生长、生化指标和肠道健康的影响, 以期为缓解鱼类热应激提供理论依据。
1. 材料与方法
1.1 实验饲料
本实验所用基础饲料以鱼粉、鸡肉粉和棉籽浓缩蛋白为主要蛋白源, 配制的粗蛋白含量为55%, 粗脂肪含量为8.5%。研究表明日粮中添加5 g/kg NaHCO3显著缓解夏季热应激对肉鸡生长性能和肝脏健康的影响[20], 因此本实验在基础饲料中添加5 g/kg NaHCO3(纯度≥98%, 上海源叶生物科技有限公司, 中国上海)制备成实验组饲料。所有原料经过超微粉碎机粉碎, 过80目筛网, 随后用木薯淀粉逐步稀释, 并与其他原料充分混合。加水搅拌均匀后, 使用双螺杆挤压机(SLP−45, 上海渔业机械设备研究所, 中国)制备粒径约为4 mm的膨化颗粒饲料, 70℃烘干后放置4℃中保存备用。饲料配方见表 1。
表 1 实验饲料配方及化学组成(%干物质)Table 1. Diet formulation and chemical composition (% dry matter)原料Ingredient 饲料Diet CON NaHCO3 鱼粉Fish meal 42.00 42.00 鸡肉粉Chicken powder 12.00 12.00 棉籽浓缩蛋白Cottonseed protein concentrate 9.00 9.00 谷朊粉Wheat flour 5.00 5.00 豆粕Soybean meal 7.00 7.00 木薯淀粉Tapioca flour 12.00 12.00 鱼油Fish oil 2.00 2.00 豆油Soybean oil 2.00 2.00 多维多矿预混物Vitamin and mineral premix1 2.00 2.00 磷酸二氢钙Ca (H2PO4)2 1.00 1.00 氯化胆碱Choline chloride 0.30 0.30 DL-蛋氨酸DL-Methionine 0.21 0.21 L-赖氨酸盐酸盐L-Lysine hydrochloride (98%) 0.36 0.36 碳酸氢钠NaHCO3 0.00 0.50 纤维素Cellulose 5.13 4.63 化学组成Chemical composition (%) 水分Moisture 7.12 6.97 粗蛋白Crude protein 54.62 55.07 粗脂肪Crude lipid 8.52 8.38 注: 1多维多矿预混物: 七水硫酸镁, 205.39 mg/kg; 七水硫酸铁, 198.57 mg/kg; 七水硫酸锰, 50.36 mg/kg; 碘化钾, 0.07 mg/kg; 五水硫酸铜, 98.30 mg/kg; 七水硫酸锌, 442.46 mg/kg; 亚硒酸钠, 0.66 mg/kg; 七水硫酸钴, 1.67 mg/kg; 维生素A, 1800 IU/kg; 维生素D3, 390 IU/kg; 维生素E, 30 mg/kg; 维生素K3, 2.8 mg/kg; 维生素B1, 6 mg/kg; 维生素B2, 6 mg/kg; 维生素B6, 6 mg/kg; 维生素B12, 0.04 mg/kg; 维生素C, 70 mg/kg; 泛酸钙, 15 mg/kg; 烟酰胺, 45 mg/kg; 叶酸, 2 mg/kg; D-生物素, 0.05 mg/kg; 肌醇, 40 mg/kg; 玉米淀粉, 4500 mg/kg。购自广东联鲲集团有限公司Note: 1Vitamin premix: Minerals premix: MgSO4.7H2O, 205.39 mg/kg; FeSO4.7H2O, 198.57 mg/kg; MnSO4.7H2O, 50.36 mg/kg; KI, 0.07 mg/kg; CuSO4.5H2O, 98.30 mg/kg; ZnSO4.7H2O, 442.46 mg/kg; Na2SeO3, 0.66 mg/kg; CoSO4.7H2O, 1.67 mg/kg; Vitamin A, 1800 IU/kg; Vitamin D3, 390 IU/kg; Vitamin E, 30 mg/kg; Vitamin K3, 2.8 mg/kg; Vitamin B1, 6 mg/kg; Vitamin B2, 6 mg/kg; Vitamin B6, 6 mg/kg; Vitamin B12, 0.04 mg/kg; Vitamin C, 70 mg/kg; Calcium pantothenate, 15 mg/kg; Nicotinamide, 45 mg/kg; Folic acid, 2 mg/kg; D-biotin, 0.05 mg/kg; Myo-inositol, 40 mg/kg; Corn starch, 4500 mg/kg. Purchased from Guangdong Liankun Group Co., Ltd (Guangdong, China) 1.2 实验鱼及养殖条件
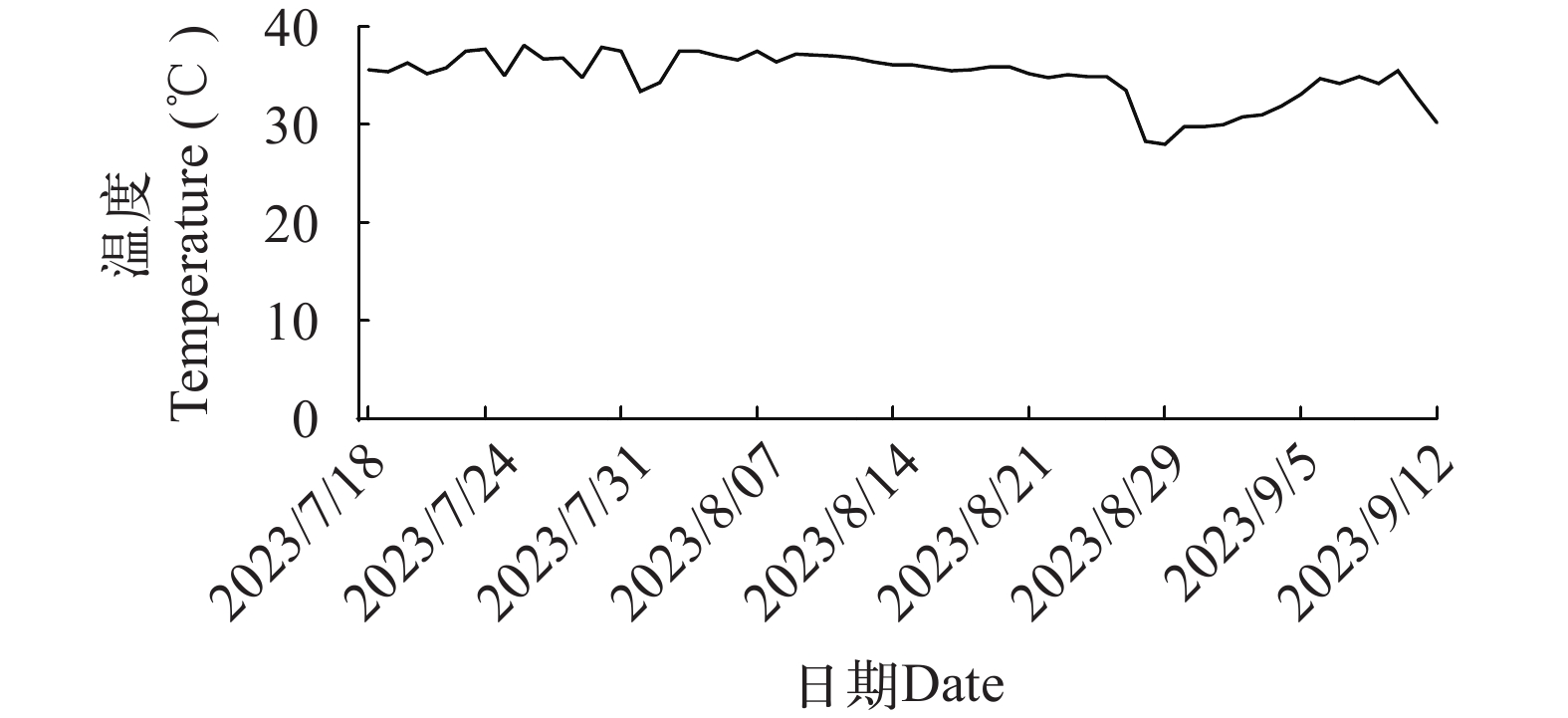
实验所用的大口黑鲈购自湖北正好水产养殖公司(中国湖北)。养殖实验在湖北省石首市老河长江四大家鱼原种场的池塘网箱(2 m×2 m×2 m)中进行。实验鱼在网箱内暂养, 使用基础饲料饲喂2周。实验鱼在正式实验前饥饿24h。挑选健康、规格均匀的420尾幼鱼[初始均重: (20.26±0.08) g], 称重后随机放入6个网箱中。实验期间, 每日饱食投喂2次(6:30、17:30)。养殖实验持续8周。实验水温为(34.9±0.3)℃(图 1), 溶解氧>6 mg/L, 氨氮含量为0.20—0.34 mg/L, 亚硝酸盐含量为0.009—0.014 mg/L, pH为6.8—7.2。
1.3 样品采集
在实验开始前, 选取规格相近的3组实验鱼作为初样, 保存在−20℃冰箱中备用。在养殖实验结束后, 使用MS-222 (60 mg/L, Sigma, 美国)对实验鱼进行麻醉, 并进行称重和计数。每个网箱随机抽取2尾鱼作为末样, 用于基本组分的测定。同时, 随机抽取2尾鱼, 使用肝素钠抗凝剂浸润过的1 mL无菌注射器采集尾部静脉血样, 4℃下离心分离血浆, 并在冰浴条件下快速取出肠道样品。血浆和肠道样品均存放于−80℃保存备用。部分肠道组织用4%多聚甲醛固定液固定, 通过组织学分析肠道形态[21]。
1.4 样品生化分析
实验饲料和鱼体基本组分(粗蛋白、粗脂肪、水分和灰分)含量按照AOAC[22]标准方法进行测定。采用全自动生化分析仪(BS-460, 深圳迈瑞生物医疗电子股份有限公司, 深圳)测定血浆总蛋白(Total protein, P/N:105-015578-00)、白蛋白(Albumin, P/N:105-000450-00)、碱性磷酸酶(Alkaline phosphatase, P/N: 105-000444-00)和葡萄糖(Glucose, P/N:105-000949-00)[23]。
迅速剪取约50 mg中肠组织, 置入1.5 mL离心管底部, 按1﹕9的比例加入0.68%的无菌生理盐水, 在低温条件下用高速组织匀浆机匀浆。随后在4℃、3000 r/min条件下离心10 min, 取上清制成10%的肠道组织匀浆液, 使用考马斯亮蓝试剂盒(碧云天生物技术研究所, P0006)测定蛋白质含量。同时, 使用南京建成生物工程研究所的试剂盒检测总抗氧化能力(T-AOC, A015-2)、谷胱甘肽过氧化物酶(GPx, A005-1-2)、过氧化氢酶(CAT, A007-1-1)、超氧化物歧化酶(SOD, A001-3-2)活性及丙二醛(MDA, A003-1-2)含量, 检测肠道胰蛋白酶(A080-2-2)、脂肪酶(A054-2-1)和α-淀粉酶(C016-1-2)活性。
1.5 基因表达分析
根据Wu等[24]的方法, 使用Trizol试剂(Invitrogen, 美国)提取肠道组织RNA, 利用M-MLV逆转录酶(M-MLV)将RNA反转录为cDNA。使用LightCycle 480 II系统(Roche, Basel, Switzerland)和2× TSINGKE® Master SYBR Green Ι qPCR Mix-UDG(Without ROX)荧光染料进行实时荧光定量PCR检测。β-actin为内参基因, 根据引物扩增效率计算目标基因的相对表达水平[25]。相关基因的引物序列见表 2。
表 2 本实验所用引物序列表Table 2. Primers used in the present experiment基因名Gene 引物序列Primer sequence(5′—3′) 登录号Accession number β-actin CTGTGGTGGTGAATGAGTAGCC PRJNA725023 CATCCTCCGTTTGGACTTGG Heat shock protein 90 (hsp90) ACCCCTCTCTCCCTGGGAATT XM_038708948.1 GCACGCTCACCCTCATAAACC Heat shock protein 70 (hsp70) GTCCTACGCCTTCAACACGA XM_038708948.1 GCTGATGGTCTCGTCACACT Heat shock protein 60 (hsp60) AAGCAGGTGAGGCCTGTGTG XM_038721348.1 AGCATGAGGGCACGAGCATC Heat shock protein 30 (hsp30) CTACGTACAGGGTTGCGGGG XM_038732984.1 ATGGACTCAGCTACATGTTGTTCT Heat shock protein 27 (hsp27) ATCGGGCAAATCGCCTTCA XM_038709260.1 AGACCACCGTGTGATTGCT Interleukin 1β (il-1β) CGTGACTGACAGCAAAAAGAGG XM_038696252.1 GATGCCCAGAGCCACAGTTC Interleukin 15 (il15) GTATGCTGCTTCTGTGCCTGG XM_038713163.1 AGCGTCAGATTTCTCAATGGTGT Interleukin 10 (il10) CGATTCTGCCAACAGCCTTG XM_038696252.1 GCTCGTCGAAGATCTGCTGT Tumor necrosis factor α (tnfα) CTTCGTCTACAGCCAGGCATCG XM_038710731.1 TTTGGCACACCGACCTCACC Transforming growth factor β (tgfβ) GCTCAAAGAGAGCGAGGATG XM_038693206.1 TCCTCTACCATTCGCAATCC Nuclear factor (erythroid-derived 2)-like 2 TCACCAAAGACAAGCGTAA XM_038720536.1 (nrf2) CAGGCAGATTGATAATCATAGA Kelch-like ECH-associated protein-1 GATAGACAGCGTGGTCAAGGC MW465394 (keap1) TGAAGAACTCCTCCTGGGTCG Catalase (cat) GTTCCCGTCCTTCATCCACT MK614708.1 CAGGCTCCAGAAGTCCCACA Superoxide dismutase 1 (sod1) GCGTGGGTAGATGGTTT XM_038713969.1 AGGGTTGATGGGCAGTA Glutathione peroxidase 1a (gpx1a) CCCTGCAATCAGTTTGGACA XM_038697220.1 TTGGTTCAAAGCCATTCCCT Zonula occludens-1 (zo-1) ATCTCAGCAGGGATTCGACG XM_038701018.1 CTTTTGCGCTGGCGTTGG Mucin 2 (muc2) AAAATAAGGAGGGTTGGG XM_038706114.1 GCTGTGCCGTGGTTCTAG Occludin GATATGGTGGCAGCTACGGT XM_038715419.1 TCCTACTGCGGACAGTGTTG Claudin CCAGGGAAGGGGAGCAATG XM_038713307.1 GCTCTTTGAACCAGTGCGAC Junctional adhesion molecule (jam) TAGGTTTGCTGCTCTTTGGTCT NM_001004667 GTCTGAATCCGTTAGCCTCATC 1.6 数据统计分析
所有数据均以平均数±标准误表示(n=6)。数据的正态分布情况通过SPSS 19.0软件中的Shapiro-Wilk检验进行评估。随后, 采用独立样本t检验(Independent sample t-test)比较对照组(CON)和碳酸氢钠(NaHCO3)组的差异。P<0.05被认为具有显著性差异。
2. 结果
2.1 生长与鱼体组成
如表 3所示, 在高温胁迫下, 饲料中添加NaHCO3显著提高了大口黑鲈的饲料效率(FE)、蛋白质效率(PER)和蛋白质沉积率(PRE; P<0.05)。与对照组相比, NaHCO3组的终末体重(FBW)、摄食量(FI)、增重率(WGR)、特定生长率(SGR)和存活率(SR)无显著差异(P>0.05)。鱼体组分分析结果显示, NaHCO3组的全鱼粗脂肪含量显著低于对照组(P<0.05)。
表 3 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈生长、饲料利用和鱼体营养成分的影响Table 3. Effects of dietary NaHCO3 supplementation on growth, feed utilization, and fish nutritional composition of largemouth bass under chronic heat stress指标Item 饲料Diet CON NaHCO3 初始体重 IBW1 (g) 20.25±0.11 20.17±0.17 终末体重 FBW2 (g) 92.93±0.88 102.73±3.44 摄食量 FI3 (g/fish) 93.40±2.76 93.01±3.17 增重率 WGR4 (%) 358.79±6.79 409.13±14.40 饲料效率 FE5 (%) 83.92±1.38a 86.27±1.16b 特定生长率 SGR6 (%/d) 2.72±0.03 2.90±0.05 存活率 SR7 (%) 86.19±0.48 87.14±3.78 蛋白质效率 PER8 1.41±0.03a 1.56±0.02b 蛋白质沉积率 PRE9 (%) 20.66±1.33a 26.18±0.39b 初始鱼体营养成分 Initial fish nutritional composition 粗蛋白 Crude protein (%) 8.10±0.14 8.10±0.14 粗脂肪 Crude lipid (%) 20.01±1.09 20.01±1.09 灰分 Ash (%) 16.56±0.52 16.56±0.52 水分 Moisture (%) 65.10±1.24 65.10±1.24 终末鱼体营养成分Final fish
nutritional composition (%)粗蛋白 Crude protein 15.77±0.82 16.67±0.21 粗脂肪 Crude lipid 7.47±0.21b 6.76±0.18a 灰分 Ash 15.65±1.85 17.42±0.35 水分 Moisture 71.45±1.29 70.52±0.86 注: 表中数据表示为平均值±标准误(n=3), 同一行中不同数值的英文字母上标表示差异显著(P<0.05)。The data are expressed as means±SE (n=3), and the different superscript letters in the same row indicate significant differences (P<0.05); 1初始体重 IBW (g): Initial body weight; 2终末体重 FBW (g): Final body weight; 3摄食量 FI (g/fish)=干物质摄食量/鱼数目; Feed intake (FI, g/fish)=dry feed intake/number of fish; 4增重率WGR (%)=100×(终末体重–初始体重)/初始体重; Weight gain rate (WGR, %)=100×(FBW–IBW)/IBW; 5饲料效率 FE(%)=(100×鱼体湿重增重量)/干物质摄食量; Feed efficiency (FE, %)=(100×fresh body weight gain)/dry feed intake; 6特定增长率SGR (%/d)=100×[ln(终末体重)–ln(初始体重)]/投喂天数; Specific grow rate (SGR, %/d)=100×[ln (final body weight)–ln (initial body weight)]/days; 7存活率SR (%)=(存活鱼总数/起始鱼总数)×100; Survival rate (SR, %)=100×(final fish number/initial fish number); 8蛋白质效率PER = (终末体重–初始体重)/蛋白摄入量; Protein efficiency ratio (PER)=(final body weight–initial body weight)/protein intake; 9蛋白质沉积率PRE (%)=100×(末样体蛋白×终末体重–初样体蛋白×初始体重)/蛋白摄入量; Protein retention efficiency (PRE, %)=100×(final body protein×final body weight–initial body protein×initial body weight)/protein intake 2.2 血浆生化指标
大口黑鲈的血浆生化指标如图 2所示。在高温胁迫下, 饲料中添加NaHCO3显著提高了大口黑鲈血浆中碱性磷酸酶和总蛋白的含量(P<0.05), 对白蛋白和葡萄糖水平无显著影响(P>0.05)。
![]() 图 2 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈血浆生化指标的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 2. Effects of dietary NaHCO3 on serum biochemical indices in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 2 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈血浆生化指标的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 2. Effects of dietary NaHCO3 on serum biochemical indices in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)2.3 肠道抗氧化及消化酶指标
如表 4所示, 与对照组相比, 饲料中添加NaHCO3显著提高了大口黑鲈肠道中的总抗氧化能力(T-AOC)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GPx)活性(P<0.05), 对丙二醛(MDA)含量无显著影响(P>0.05)。在肠道消化酶活性方面, NaHCO3显著提高了肠道淀粉酶和胰蛋白酶的活性(P<0.05), 但对脂肪酶活性无显著影响(P>0.05)。
表 4 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈肠道抗氧化能力和消化酶活性的影响Table 4. Effects of dietary NaHCO3 on antioxidant enzyme activity and digestive enzymes in gut of largemouth bass under chronic heat stress指标Item CON NaHCO3 总抗氧化能力T-AOC
(nmol/mg prot)0.31±0.01a 0.38±0.02b 超氧化物歧化酶SOD
(U/mg prot)15.88±2.83a 31.32±2.85b 丙二醛MDA
(nmol/mg prot)2.62±0.15 2.31±0.40 过氧化氢酶CAT
(U/mg prot)13.50±1.84a 25.71±1.75b 谷胱甘肽过氧化物酶GPx
(U/mg prot)10.45±0.67a 27.69±2.30b 脂肪酶 (U/g prot) 1.73±0.17 2.65±0.41 α-淀粉酶 (U/mg prot) 0.88±0.08a 1.37±0.06b 胰蛋白酶 (U/mg prot) 2004.08±619.86a 6322.22±1502.28b 注: 同一行不同上标字母表示差异显著(P<0.05)Note: Different superscript letters in the same row indicate significant differences (P<0.05) 2.4 肠道抗氧化、炎症和热休克蛋白家族相关基因表达
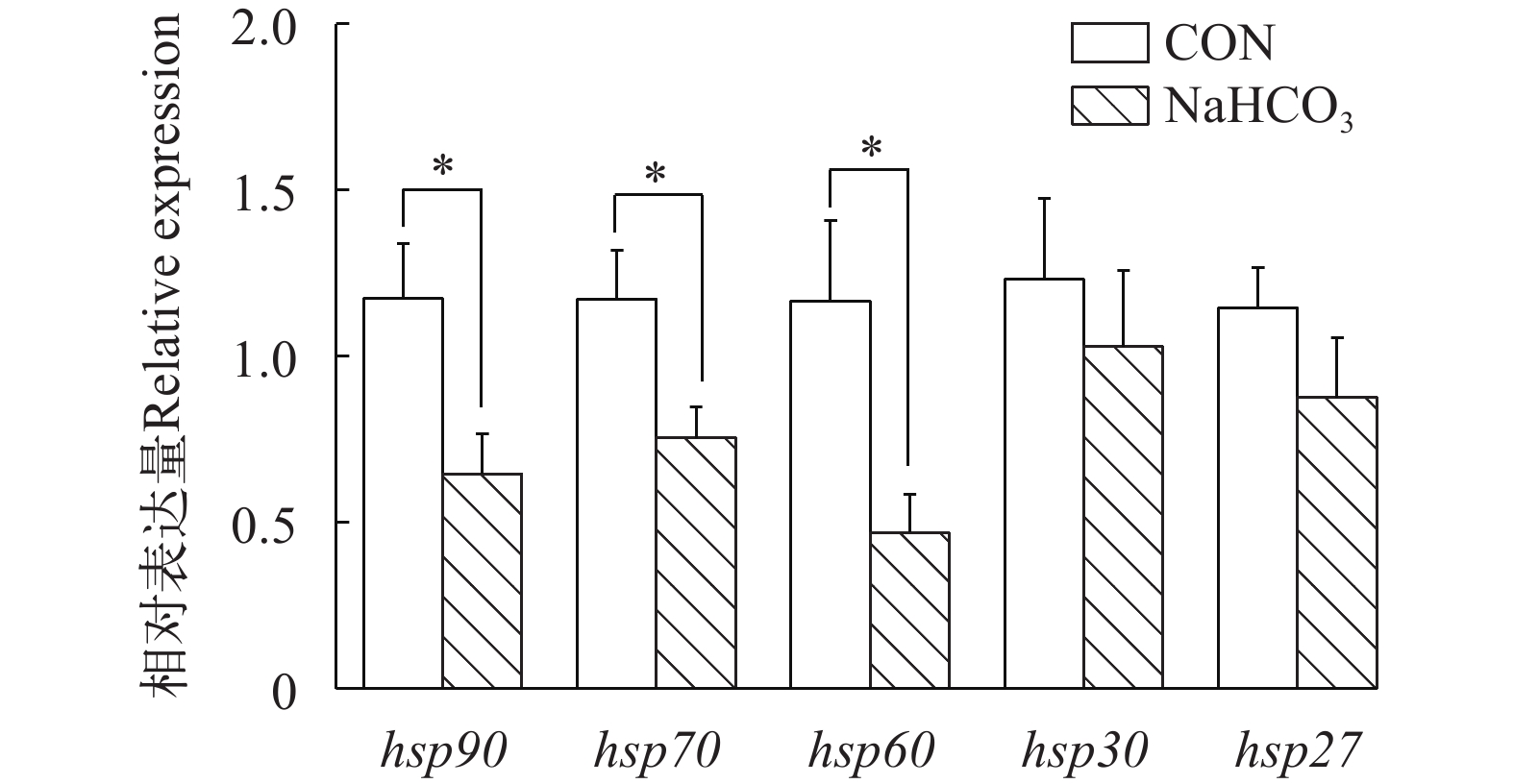
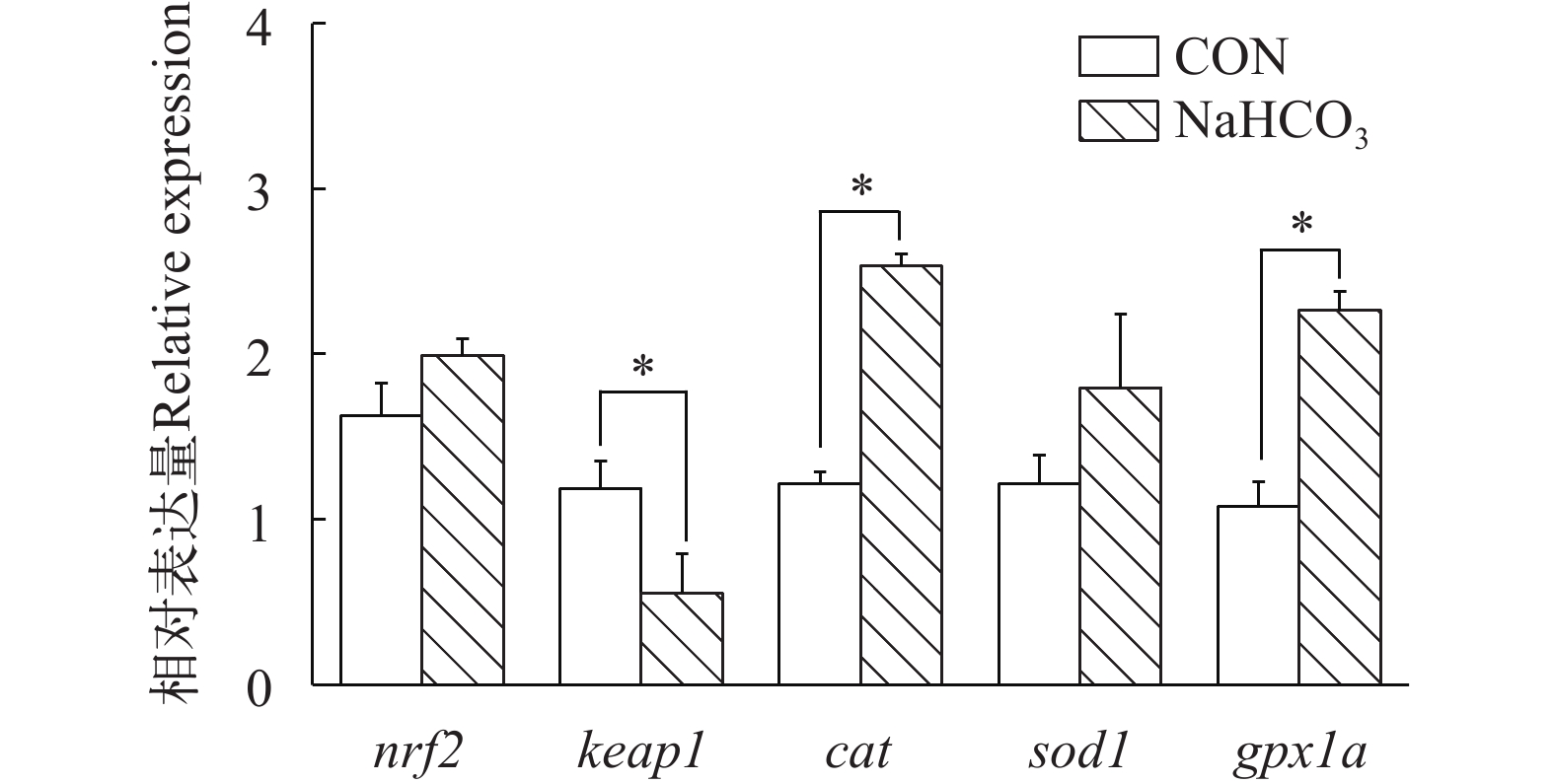
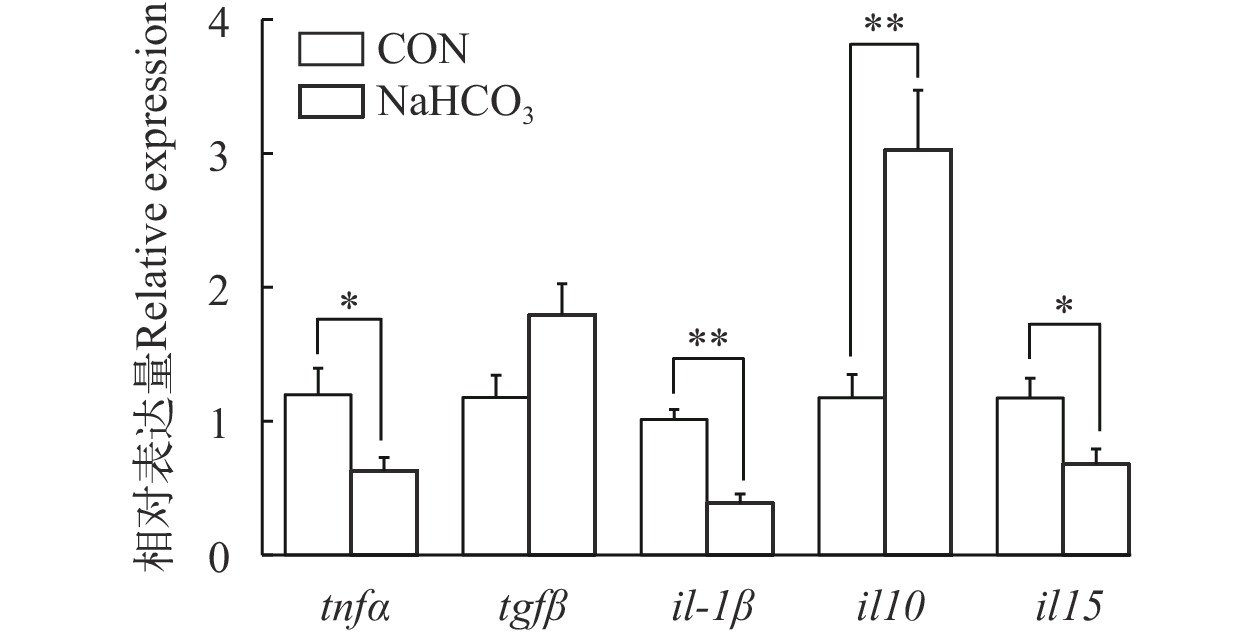
在高温胁迫条件下, 与对照组相比, 饲料中添加NaHCO3显著降低了肠道热休克蛋白(hsp90、hsp70、hsp60)的基因表达(P<0.05), 而对hsp30和hsp27的转录水平无显著影响(P>0.05; 图 3)。如图 4所示, NaHCO3处理显著上调了肠道抗氧化相关基因(cat、gpx1a)的表达, 并显著下调了keap1的基因表达(P<0.05), 但是不影响nrf2和sod1的基因表达(P>0.05)。在慢性高温胁迫条件下, 添加NaHCO3显著下调了促炎症细胞因子(il-1β、tnfα和il15)的基因表达, 同时上调了抗炎细胞因子il10的表达(P<0.05; 图 5)。
![]() 图 3 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道热休克蛋白相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 3. Effect of NaHCO3 on heat shock protein-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 3 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道热休克蛋白相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 3. Effect of NaHCO3 on heat shock protein-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)![]() 图 4 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道抗氧化相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 4. Effect of NaHCO3 on antioxidant-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 4 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道抗氧化相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 4. Effect of NaHCO3 on antioxidant-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)![]() 图 5 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道炎症相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 5. Effect of NaHCO3 on inflammation-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 5 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道炎症相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 5. Effect of NaHCO3 on inflammation-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)2.5 肠道组织学及紧密连接相关基因的表达
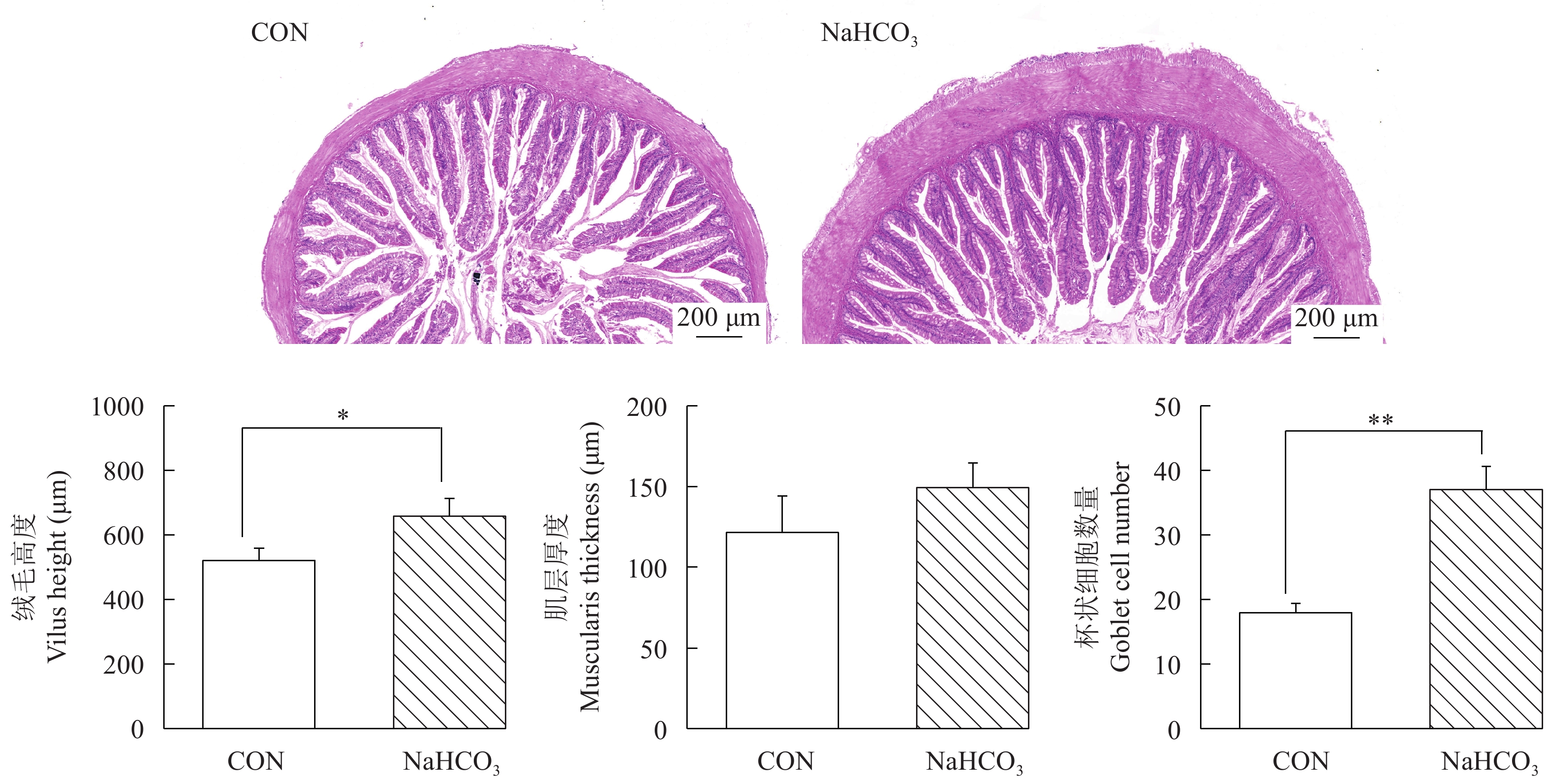
NaHCO3添加对慢性高温胁迫下大口黑鲈肠道组织学的影响如图 6所示。HE染色结果显示, 在慢性高温胁迫8周后, 对照组大口黑鲈肠道绒毛明显缩短且排列稀疏, 部分黏膜细胞脱落; 而NaHCO3组肠道形态较为完整, 结构清晰, 肠绒毛排列紧密。两组间肌层厚度差异不显著, 但NaHCO3显著提高了大口黑鲈肠道绒毛长度和杯状细胞数量。同时, NaHCO3处理显著提高了慢性高温胁迫条件下大口黑鲈肠道组织中紧密连接相关基因zo1、jam、occludin和muc2的表达水平(P<0.05; 图 7)。
![]() 图 6 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道组织学的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 6. Effects of NaHCO3 on gut histology in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 6 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道组织学的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 6. Effects of NaHCO3 on gut histology in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)![]() 图 7 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道紧密连接相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 7. Effects of NaHCO3 on intestinal tight junction-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
图 7 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道紧密连接相关基因表达的影响柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示Figure 7. Effects of NaHCO3 on intestinal tight junction-related gene expression in largemouth bass under chronic heat stressThe * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)3. 讨论
3.1 NaHCO3对慢性高温胁迫下大口黑鲈生长性能和血浆生化指标的影响
水温是水产养殖的重要因素, 高温会引起水产养殖动物的应激, 从而影响生长和免疫等。在中华绒螯蟹(Eriocheir sinensis)中, 高温应激增加了中华绒螯蟹摄食量, 并促进其生长[26]。饲料中添加鱼腥草可改善高温应激对大口黑鲈的影响, 提高其耐高温能力[5]。碳酸氢钠在畜禽养殖中广泛应用于缓解高温应激, 不仅能够改善畜禽的抗应激能力, 还能提高饲料利用率并加速营养物质的吸收[27]。饲料中添加NaHCO3可缓解热应激对山羊的饲料消耗、消化能力、代谢和生理功能的不利影响, 从而提高雄鹿的生长性能和饲料利用[19]。在本实验中, 饲料中添加NaHCO3对高温胁迫下的大口黑鲈的生长性能无显著影响, 但提高了饲料利用率。在合方鲫(Carassius auratus)中, 其增重率和特定生长率随着NaHCO3浓度的增加反而下降[28]。因此, NaHCO3的作用可能与品种、胁迫时间、应激强度和养殖环境有关。
血浆总蛋白含量常被用来反映鱼类的免疫水平[29]。结果显示, 饲料中添加NaHCO3显著提高了大口黑鲈血浆中总蛋白含量, 对白蛋白无显著影响, 这与之前的研究一致[30]。血浆碱性磷酸酶是一种在鱼类等生物体内广泛存在的酶, 其活性通常与机体的营养状况、免疫及代谢状态等密切相关[31]。碱性磷酸酶活性升高不仅可以增强机体的免疫力, 还可以通过调节机体的能量代谢来适应高温胁迫。在本实验中, NaHCO3组的血浆碱性磷酸酶活性显著高于对照组。这可能是因为NaHCO3能够中和部分酸性代谢产物, 维持机体酸碱平衡, 从而减轻热应激对机体的损伤。
3.2 NaHCO3对慢性高温胁迫下大口黑鲈肠道抗氧化能力的影响
氧化应激主要是活性氧(ROS)和抗氧化能力之间的不平衡引起的, 而热应激是导致活性代谢物清除失衡的主要应激源之一[32]。总抗氧化能力(T-AOC)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和谷胱甘肽过氧化物酶(GSH-Px)是机体抗氧化防御系统的重要指标, 能有效清除体内ROS, 减轻脂质过氧化反应, 抵御氧化应激[33]。与肉鸡中的研究结果一致[27], NaHCO3添加后通过增加大口黑鲈肠道抗氧化酶活性来减轻慢性高温胁迫引起的氧化应激。同样, 在H9C2细胞中, 维生素C和NaHCO3联合使用显著提高了热应激抑制的抗氧化酶活性[13]。Nrf2作为一种核内转录因子, 通过调控一系列抗氧化相关基因的表达, 协同调控ROS引起的氧化应激[34]。本实验中NaHCO3处理显著上调了大口黑鲈肠道抗氧化基因(cat、gpx1a)的表达, 并显著下调了keap1的基因表达。在杂交鲟(Acipenser baerii ♀×A. schrenkii ♂)[35]和梭鲈(Sander lucioperca)[33]中也观察到类似的结果。这些结果表明, NaHCO3添加后通过增强大口黑鲈抗氧化能力来缓解高温胁迫引起的氧化损伤。
3.3 NaHCO3对慢性高温胁迫下大口黑鲈肠道热休克蛋白和炎症基因表达的影响
在热应激条件下, 热休克蛋白(HSP)通过不同的机制和作用方式共同维护细胞的正常生理功能和稳定性,被认为是细胞抵抗损伤和适应环境应激能力的重要指标[36]。Hsp90是HSP家族中分布最广泛的成员, 具有维持细胞结构完整性、重新折叠异常折叠的蛋白质和提高细胞耐热性等作用[26]。在中华绒螯蟹[26]、虹鳟(Oncorhynchus mykiss)[37]和尼罗罗非鱼(Oreochromis niloticus)[38]中, 高温胁迫增加了hsp90的基因表达。本实验中, 与对照组相比, 饲料中添加NaHCO3显著降低了大口黑鲈肠道热休克蛋白(hsp60、hsp70和hsp90)的基因表达, 表明NaHCO3缓解了高温胁迫引起的肠道损伤[13]。研究表明, 过高的温度刺激则会影响尼罗罗非鱼肠道的免疫系统[39]。高温胁迫上调了虹鳟[37]、杂交鲟[8]和尼罗罗非鱼[39]肠道抗炎因子的表达, 引起了肠道氧化损伤。在慢性高温胁迫下, 添加NaHCO3显著下调了大口黑鲈促炎症细胞因子(il-1β、tnfα和il15)的基因表达, 上调了抗炎细胞因子il10的表达, 表明NaHCO3可能具有抗炎作用。
3.4 NaHCO3对慢性高温胁迫下大口黑鲈肠道组织学及紧密连接相关基因表达的影响
高温胁迫可能会通过影响鱼类消化酶的活性, 改变肠道吸收能力, 使其对食物的消化能力下降[28, 40, 41]。在本实验中, NaHCO3添加后显著提高了大口黑鲈肠道淀粉酶和胰蛋白酶的活性, 可能是提高饲料效率的原因之一。肠道屏障是机体抵御外来入侵的关键。肠黏膜屏障损伤后, 外界环境中的有害病原菌及外来毒素可侵入肠黏膜, 进而对宿主的免疫功能产生不利影响[42]。肠道黏膜层表面的褶皱和微绒毛, 在营养物质的消化和吸收中发挥着关键的作用。热应激胁迫下, 虹鳟肠道绒毛高度、肌层厚度以及杯状细胞数量均显著减少[37]。与对照组相比, 慢性高温胁迫8周后, 大口黑鲈肠道受损, 部分黏膜细胞脱落。而在饲料中添加了NaHCO3后则改善了肠道组织学形态, 增加了绒毛高度、肌层厚度以及杯状细胞数量。紧密连接蛋白主要由外周膜蛋白ZO-1和跨膜蛋白OCLN和claudins组成, 是肠黏膜屏障的重要组成部分, 在维持细胞通透性和屏障功能方面发挥着关键作用[43]。因此, 紧密连接蛋白的下调将对肠道结构和屏障功能产生不利影响。在虹鳟中, 热应激显著下调了肠道中ZO-1、occludin和claudin-8的基因表达[37]。慢性高温胁迫下, 大口黑鲈肠道zo1、jam、occludin、muc2的表达水平显著增加。结合肠道形态学结果表明饲料中添加NaHCO3有效缓解大口黑鲈高温应激引起的肠道屏障损伤, 从而提高慢性高温胁迫下大口黑鲈的肠道健康。
4. 结论
综上所述, 本研究探讨了饲料中添加5 g/kg的NaHCO3对慢性高温应激引起的大口黑鲈肠道损伤的缓解作用。结果表明, 在慢性高温应激下, 饲料中添加NaHCO3通过改善肠道屏障功能、减轻肠道氧化应激和炎症反应, 有效缓解了高温诱导的大口黑鲈肠道损伤, 从而提高了大口黑鲈的饲料利用。本研究为改善鱼类慢性高温胁迫下的肠道健康提供了理论依据。
-
![]()
图 2 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈血浆生化指标的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 2. Effects of dietary NaHCO3 on serum biochemical indices in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
![]()
图 3 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道热休克蛋白相关基因表达的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 3. Effect of NaHCO3 on heat shock protein-related gene expression in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
![]()
图 4 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道抗氧化相关基因表达的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 4. Effect of NaHCO3 on antioxidant-related gene expression in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
![]()
图 5 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道炎症相关基因表达的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 5. Effect of NaHCO3 on inflammation-related gene expression in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
![]()
图 6 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道组织学的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 6. Effects of NaHCO3 on gut histology in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
![]()
图 7 NaHCO3添加对慢性高温胁迫下大口黑鲈肠道紧密连接相关基因表达的影响
柱状图上方的*或者**表示各处理间的差异, *表示显著性差异(P<0.05), **表示极显著差异(P<0.01); 结果用均值±标准误(n=6)表示
Figure 7. Effects of NaHCO3 on intestinal tight junction-related gene expression in largemouth bass under chronic heat stress
The * or ** at the top of the bar chart indicates significant differences between treatments; * indicates a significant difference (P<0.05), and ** indicates an extremely significant difference (P<0.01); The values are expressed as the mean±SEM (n=6)
表 1 实验饲料配方及化学组成(%干物质)
Table 1 Diet formulation and chemical composition (% dry matter)
原料Ingredient 饲料Diet CON NaHCO3 鱼粉Fish meal 42.00 42.00 鸡肉粉Chicken powder 12.00 12.00 棉籽浓缩蛋白Cottonseed protein concentrate 9.00 9.00 谷朊粉Wheat flour 5.00 5.00 豆粕Soybean meal 7.00 7.00 木薯淀粉Tapioca flour 12.00 12.00 鱼油Fish oil 2.00 2.00 豆油Soybean oil 2.00 2.00 多维多矿预混物Vitamin and mineral premix1 2.00 2.00 磷酸二氢钙Ca (H2PO4)2 1.00 1.00 氯化胆碱Choline chloride 0.30 0.30 DL-蛋氨酸DL-Methionine 0.21 0.21 L-赖氨酸盐酸盐L-Lysine hydrochloride (98%) 0.36 0.36 碳酸氢钠NaHCO3 0.00 0.50 纤维素Cellulose 5.13 4.63 化学组成Chemical composition (%) 水分Moisture 7.12 6.97 粗蛋白Crude protein 54.62 55.07 粗脂肪Crude lipid 8.52 8.38 注: 1多维多矿预混物: 七水硫酸镁, 205.39 mg/kg; 七水硫酸铁, 198.57 mg/kg; 七水硫酸锰, 50.36 mg/kg; 碘化钾, 0.07 mg/kg; 五水硫酸铜, 98.30 mg/kg; 七水硫酸锌, 442.46 mg/kg; 亚硒酸钠, 0.66 mg/kg; 七水硫酸钴, 1.67 mg/kg; 维生素A, 1800 IU/kg; 维生素D3, 390 IU/kg; 维生素E, 30 mg/kg; 维生素K3, 2.8 mg/kg; 维生素B1, 6 mg/kg; 维生素B2, 6 mg/kg; 维生素B6, 6 mg/kg; 维生素B12, 0.04 mg/kg; 维生素C, 70 mg/kg; 泛酸钙, 15 mg/kg; 烟酰胺, 45 mg/kg; 叶酸, 2 mg/kg; D-生物素, 0.05 mg/kg; 肌醇, 40 mg/kg; 玉米淀粉, 4500 mg/kg。购自广东联鲲集团有限公司Note: 1Vitamin premix: Minerals premix: MgSO4.7H2O, 205.39 mg/kg; FeSO4.7H2O, 198.57 mg/kg; MnSO4.7H2O, 50.36 mg/kg; KI, 0.07 mg/kg; CuSO4.5H2O, 98.30 mg/kg; ZnSO4.7H2O, 442.46 mg/kg; Na2SeO3, 0.66 mg/kg; CoSO4.7H2O, 1.67 mg/kg; Vitamin A, 1800 IU/kg; Vitamin D3, 390 IU/kg; Vitamin E, 30 mg/kg; Vitamin K3, 2.8 mg/kg; Vitamin B1, 6 mg/kg; Vitamin B2, 6 mg/kg; Vitamin B6, 6 mg/kg; Vitamin B12, 0.04 mg/kg; Vitamin C, 70 mg/kg; Calcium pantothenate, 15 mg/kg; Nicotinamide, 45 mg/kg; Folic acid, 2 mg/kg; D-biotin, 0.05 mg/kg; Myo-inositol, 40 mg/kg; Corn starch, 4500 mg/kg. Purchased from Guangdong Liankun Group Co., Ltd (Guangdong, China)  下载: 导出CSV
下载: 导出CSV
表 2 本实验所用引物序列表
Table 2 Primers used in the present experiment
基因名Gene 引物序列Primer sequence(5′—3′) 登录号Accession number β-actin CTGTGGTGGTGAATGAGTAGCC PRJNA725023 CATCCTCCGTTTGGACTTGG Heat shock protein 90 (hsp90) ACCCCTCTCTCCCTGGGAATT XM_038708948.1 GCACGCTCACCCTCATAAACC Heat shock protein 70 (hsp70) GTCCTACGCCTTCAACACGA XM_038708948.1 GCTGATGGTCTCGTCACACT Heat shock protein 60 (hsp60) AAGCAGGTGAGGCCTGTGTG XM_038721348.1 AGCATGAGGGCACGAGCATC Heat shock protein 30 (hsp30) CTACGTACAGGGTTGCGGGG XM_038732984.1 ATGGACTCAGCTACATGTTGTTCT Heat shock protein 27 (hsp27) ATCGGGCAAATCGCCTTCA XM_038709260.1 AGACCACCGTGTGATTGCT Interleukin 1β (il-1β) CGTGACTGACAGCAAAAAGAGG XM_038696252.1 GATGCCCAGAGCCACAGTTC Interleukin 15 (il15) GTATGCTGCTTCTGTGCCTGG XM_038713163.1 AGCGTCAGATTTCTCAATGGTGT Interleukin 10 (il10) CGATTCTGCCAACAGCCTTG XM_038696252.1 GCTCGTCGAAGATCTGCTGT Tumor necrosis factor α (tnfα) CTTCGTCTACAGCCAGGCATCG XM_038710731.1 TTTGGCACACCGACCTCACC Transforming growth factor β (tgfβ) GCTCAAAGAGAGCGAGGATG XM_038693206.1 TCCTCTACCATTCGCAATCC Nuclear factor (erythroid-derived 2)-like 2 TCACCAAAGACAAGCGTAA XM_038720536.1 (nrf2) CAGGCAGATTGATAATCATAGA Kelch-like ECH-associated protein-1 GATAGACAGCGTGGTCAAGGC MW465394 (keap1) TGAAGAACTCCTCCTGGGTCG Catalase (cat) GTTCCCGTCCTTCATCCACT MK614708.1 CAGGCTCCAGAAGTCCCACA Superoxide dismutase 1 (sod1) GCGTGGGTAGATGGTTT XM_038713969.1 AGGGTTGATGGGCAGTA Glutathione peroxidase 1a (gpx1a) CCCTGCAATCAGTTTGGACA XM_038697220.1 TTGGTTCAAAGCCATTCCCT Zonula occludens-1 (zo-1) ATCTCAGCAGGGATTCGACG XM_038701018.1 CTTTTGCGCTGGCGTTGG Mucin 2 (muc2) AAAATAAGGAGGGTTGGG XM_038706114.1 GCTGTGCCGTGGTTCTAG Occludin GATATGGTGGCAGCTACGGT XM_038715419.1 TCCTACTGCGGACAGTGTTG Claudin CCAGGGAAGGGGAGCAATG XM_038713307.1 GCTCTTTGAACCAGTGCGAC Junctional adhesion molecule (jam) TAGGTTTGCTGCTCTTTGGTCT NM_001004667 GTCTGAATCCGTTAGCCTCATC
下载: 导出CSV
表 3 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈生长、饲料利用和鱼体营养成分的影响
Table 3 Effects of dietary NaHCO3 supplementation on growth, feed utilization, and fish nutritional composition of largemouth bass under chronic heat stress
指标Item 饲料Diet CON NaHCO3 初始体重 IBW1 (g) 20.25±0.11 20.17±0.17 终末体重 FBW2 (g) 92.93±0.88 102.73±3.44 摄食量 FI3 (g/fish) 93.40±2.76 93.01±3.17 增重率 WGR4 (%) 358.79±6.79 409.13±14.40 饲料效率 FE5 (%) 83.92±1.38a 86.27±1.16b 特定生长率 SGR6 (%/d) 2.72±0.03 2.90±0.05 存活率 SR7 (%) 86.19±0.48 87.14±3.78 蛋白质效率 PER8 1.41±0.03a 1.56±0.02b 蛋白质沉积率 PRE9 (%) 20.66±1.33a 26.18±0.39b 初始鱼体营养成分 Initial fish nutritional composition 粗蛋白 Crude protein (%) 8.10±0.14 8.10±0.14 粗脂肪 Crude lipid (%) 20.01±1.09 20.01±1.09 灰分 Ash (%) 16.56±0.52 16.56±0.52 水分 Moisture (%) 65.10±1.24 65.10±1.24 终末鱼体营养成分Final fish
nutritional composition (%)粗蛋白 Crude protein 15.77±0.82 16.67±0.21 粗脂肪 Crude lipid 7.47±0.21b 6.76±0.18a 灰分 Ash 15.65±1.85 17.42±0.35 水分 Moisture 71.45±1.29 70.52±0.86 注: 表中数据表示为平均值±标准误(n=3), 同一行中不同数值的英文字母上标表示差异显著(P<0.05)。The data are expressed as means±SE (n=3), and the different superscript letters in the same row indicate significant differences (P<0.05); 1初始体重 IBW (g): Initial body weight; 2终末体重 FBW (g): Final body weight; 3摄食量 FI (g/fish)=干物质摄食量/鱼数目; Feed intake (FI, g/fish)=dry feed intake/number of fish; 4增重率WGR (%)=100×(终末体重–初始体重)/初始体重; Weight gain rate (WGR, %)=100×(FBW–IBW)/IBW; 5饲料效率 FE(%)=(100×鱼体湿重增重量)/干物质摄食量; Feed efficiency (FE, %)=(100×fresh body weight gain)/dry feed intake; 6特定增长率SGR (%/d)=100×[ln(终末体重)–ln(初始体重)]/投喂天数; Specific grow rate (SGR, %/d)=100×[ln (final body weight)–ln (initial body weight)]/days; 7存活率SR (%)=(存活鱼总数/起始鱼总数)×100; Survival rate (SR, %)=100×(final fish number/initial fish number); 8蛋白质效率PER = (终末体重–初始体重)/蛋白摄入量; Protein efficiency ratio (PER)=(final body weight–initial body weight)/protein intake; 9蛋白质沉积率PRE (%)=100×(末样体蛋白×终末体重–初样体蛋白×初始体重)/蛋白摄入量; Protein retention efficiency (PRE, %)=100×(final body protein×final body weight–initial body protein×initial body weight)/protein intake
下载: 导出CSV
表 4 饲料中添加NaHCO3对慢性高温胁迫下大口黑鲈肠道抗氧化能力和消化酶活性的影响
Table 4 Effects of dietary NaHCO3 on antioxidant enzyme activity and digestive enzymes in gut of largemouth bass under chronic heat stress
指标Item CON NaHCO3 总抗氧化能力T-AOC
(nmol/mg prot)0.31±0.01a 0.38±0.02b 超氧化物歧化酶SOD
(U/mg prot)15.88±2.83a 31.32±2.85b 丙二醛MDA
(nmol/mg prot)2.62±0.15 2.31±0.40 过氧化氢酶CAT
(U/mg prot)13.50±1.84a 25.71±1.75b 谷胱甘肽过氧化物酶GPx
(U/mg prot)10.45±0.67a 27.69±2.30b 脂肪酶 (U/g prot) 1.73±0.17 2.65±0.41 α-淀粉酶 (U/mg prot) 0.88±0.08a 1.37±0.06b 胰蛋白酶 (U/mg prot) 2004.08±619.86a 6322.22±1502.28b 注: 同一行不同上标字母表示差异显著(P<0.05)Note: Different superscript letters in the same row indicate significant differences (P<0.05)
下载: 导出CSV
-
[1] 黄天晴, 董福霖, 刘恩慧, 等. 热应激对鱼类生命活动影响的研究进展 [J]. 水产学杂志, 2023, 36(2): 109-113.] Huang T Q, Dong F L, Liu E H, et al. Research progress on effects of heat stress on fish life activities: a review [J]. Chinese Journal of Fisheries, 2023, 36(2): 109-113. [
[2] Wang Z, Luo H, Yang S. Different mechanisms for the extremely hot central-eastern China in July–August 2022 from a Eurasian large-scale circulation perspective [J]. Environmental Research Letters, 2023, 18(2): 024023. doi: 10.1088/1748-9326/acb3e5
[3] Alfonso S, Gesto M, Sadoul B. Temperature increase and its effects on fish stress physiology in the context of global warming [J]. Journal of Fish Biology, 2021, 98(6): 1496-1508. doi: 10.1111/jfb.14599
[4] 刘伟, 郑天伦, 孟庆辉, 等. 温度对大口黑鲈的影响及养殖生产中的应用 [J]. 水产养殖, 2021, 42(4): 56-57.] doi: 10.3969/j.issn.1004-2091.2021.04.015 Liu W, Zheng T L, Meng Q H, et al. Effect of temperature on largemouth bass and its application in culture and production [J]. Journal of Aquaculture, 2021, 42(4): 56-57. [ doi: 10.3969/j.issn.1004-2091.2021.04.015
[5] 薛小翠, 李彩莹, 赵建华, 等. 鱼腥草对加州鲈生长、抗氧化的影响及其缓解高温应激作用研究 [J]. 饲料工业, 2023, 44(4): 79-84.] Xue X C, Li C Y, Zhao J H, et al. Effects of Houttuynia cordata on growth performance and antioxidation of Micropterus salmoides and alleviating high temperature stress [J]. Feed Industry, 2023, 44(4): 79-84. [
[6] 黄太寿, 白俊杰, 李胜杰, 等. 我国加州鲈养殖现状和绿色发展问题探讨 [J]. 中国水产, 2017(12): 44-47.] Huang T S, Bai J J, Li S J, et al. Discussion on the current situation and green development of California bass culture in China [J]. China Fisheries, 2017(12): 44-47. [
[7] Xv Z C, Chen S X, Song G L, et al. Biochemical, histological and transcriptomic analyses for the immunological organs provide insights into heat stress-induced disease susceptibility in largemouth bass [J]. Science of the Total Environment, 2024, 912: 168758. doi: 10.1016/j.scitotenv.2023.168758
[8] Yang S, Zhang C, Xu W, et al. Heat stress decreases intestinal physiological function and facilitates the proliferation of harmful intestinal microbiota in sturgeons [J]. Frontiers in Microbiology, 2022, 13: 755369. doi: 10.3389/fmicb.2022.755369
[9] Yan D, Long X, Zhang X, et al. Identification and characterization of long non-coding RNAs in intestinal immune regulation of largemouth bass, Micropterus salmoides, under acute heat stress [J]. Comparative Biochemistry and Physiology. Part D: Genomics and Proteomics, 2023, 48: 101132. doi: 10.1016/j.cbd.2023.101132
[10] Wei D, Zhu L, Wang Y, et al. Variation in the intestinal bacterial community composition under different water temperature culture conditions in largemouth bass (Micropterus salmoides) [J]. Journal of Applied Microbiology, 2024, 135(11): lxae283. doi: 10.1093/jambio/lxae283
[11] Yu J, Zhong D, Li S, et al. Acute temperature stresses trigger liver transcriptome and microbial community remodeling in largemouth bass (Micropterus salmoides) [J]. Aquaculture, 2023, 573: 739573. doi: 10.1016/j.aquaculture.2023.739573
[12] 孟繁伊. 从第九届世界华人鱼虾营养学术研讨会看我国水产动物营养与饲料发展的现状与未来 [J]. 饲料工业, 2014, 35(4): 54-57.] Meng F Y. From the ninth symposium of world's Chinese scientists on nutrition and feeding of finfish and shellfish to see the present status and the future direction of aquaculture nutrition and feed in China [J]. Feed Industry, 2014, 35(4): 54-57. [
[13] Yin B, Tang S, Sun J, et al. Vitamin C and sodium bicarbonate enhance the antioxidant ability of H9C2 cells and induce HSPs to relieve heat stress [J]. Cell Stress and Chaperones, 2018, 23(4): 735-748. doi: 10.1007/s12192-018-0885-2
[14] Bottje W G, Harrison P C. Effect of carbonated water on growth performance of cockerels subjected to constant and cyclic heat stress temperatures [J]. Poultry Science, 1985, 64(7): 1285-1292. doi: 10.3382/ps.0641285
[15] Teeter R G, Smith M O, Owens F N, et al. Chronic heat stress and respiratory alkalosis: occurrence and treatment in broiler chicks [J]. Poultry Science, 1985, 64(6): 1060-1064. doi: 10.3382/ps.0641060
[16] Fujii T, Udy A, Licari E, et al. Sodium bicarbonate therapy for critically ill patients with metabolic acidosis: a scoping and a systematic review [J]. Journal of Critical Care, 2019, 51: 184-191. doi: 10.1016/j.jcrc.2019.02.027
[17] Kawas J R, García-Castillo R, Garza-Cazares F, et al. Effects of sodium bicarbonate and yeast on productive performance and carcass characteristics of light-weight lambs fed finishing diets [J]. Small Ruminant Research, 2007, 67(2-3): 157-163. doi: 10.1016/j.smallrumres.2005.09.011
[18] Wedekind K J, Muntifering R B, Barker K B. Effects of diet concentrate level and sodium bicarbonate on site and extent of forage fiber digestion in the gastrointestinal tract of wethers [J]. Journal of Animal Science, 1986, 62(5): 1388-1395. doi: 10.2527/jas1986.6251388x
[19] Jamal A, Rashid M A, Malik M I. Effects of sodium bicarbonate and chromium propionate supplementation on growth performance, blood indices of Beetal Bucks under heat stress [J]. Tropical Animal Health and Production, 2021, 53(5): 496. doi: 10.1007/s11250-021-02931-9
[20] Saker O A, El-Dakroury M F, Al-Sokary E T, et al. Ameliorative effect of dietary acetylsalicylic acid and sodium bicarbonate supplementation on growth and health status of broiler chicks exposed to heat-stress [J]. Alexandria Journal of Veterinary Sciences, 2020, 64(1): 54-62. doi: 10.5455/ajvs.77277
[21] 潘杰, 李雅萍, 陈琪, 等. 菜粕替代鱼粉对大鳞副泥鳅生长性能、抗氧化能力和肠道形态的影响 [J]. 水生生物学报, 2025, 1-12.] Pan J, Li Y P, Chen J, et al. Rapeseed meal replacement for fishmeal on growth performance, antioxidant capacity, and intestinal morphology of Paramisgurnus dabryanus [J]. Acta Hydrobiologica Sinica, 2025, 1-12. [
[22] Baur F J, Ensminger L G. The association of official analytical chemists (AOAC) [J]. Journal of the American Oil Chemists’ Society, 1977, 54(4): 171-172. doi: 10.1007/BF02670789
[23] 罗涵, 刘昊昆, 杨云霞, 等. 不同投喂频率下复合非粮蛋白源替代鱼粉饲料对大口黑鲈生长性能和健康的影响 [J]. 水生生物学报, 2024, 48(1): 63-76.] Luo H, Liu H K, Yang Y X, et al. Different dietary replacements of fishmeal by non-food proteins blend on growth performance, health of largemouth bass (Micropterus salmoides) at different feeding frequencies [J]. Acta Hydrobiologica Sinica, 2024, 48(1): 63-76. [
[24] Wu L, Li H, Xu W, et al. Dissimilar regulation of glucose and lipid metabolism by leptin in two strains of gibel carp (Carassius gibelio) [J]. The British Journal of Nutrition, 2021, 125(11): 1215-1229. doi: 10.1017/S0007114520003608
[25] Pfaffl M W. A new mathematical model for relative quantification in real-time RT-PCR [J]. Nucleic Acids Research, 2001, 29(9): e45. doi: 10.1093/nar/29.9.e45
[26] Liu J, Zhang C, Wang X, et al. Dietary methionine level impacts the growth, nutrient metabolism, antioxidant capacity and immunity of the Chinese mitten crab (Eriocheir sinensis) under chronic heat stress [J]. Antioxidants, 2023, 12(1): 209. doi: 10.3390/antiox12010209
[27] Selle P H, Liu S Y, Cai J, et al. Graded inclusions of sodium metabisulphite in Sorghum-based diets: I. Reduction of disulphide cross-linkages in vitro and enhancement of energy utilisation and feed conversion efficiency in broiler chickens [J]. Animal Feed Science and Technology, 2014, 190: 59-67. doi: 10.1016/j.anifeedsci.2013.12.015
[28] Lei X Y, Cao X, Sun J, et al. Effect of different concentrations of carbonate on growth performance, intestinal health and hepatic lipid metabolism of Crucian carp [J]. Aquaculture, 2024, 589: 740990. doi: 10.1016/j.aquaculture.2024.740990
[29] Zhu L Y, Nie L, Zhu G, et al. Advances in research of fish immune-relevant genes: a comparative overview of innate and adaptive immunity in teleosts [J]. Developmental & Comparative Immunology, 2013, 39(1/2): 39-62.
[30] Magouz F I, Amer A A, Faisal A, et al. The effects of dietary oregano essential oil on the growth performance, intestinal health, immune, and antioxidative responses of Nile Tilapia under acute heat stress [J]. Aquaculture, 2022, 548: 737632. doi: 10.1016/j.aquaculture.2021.737632
[31] Lallès J P. Intestinal alkaline phosphatase in the gastrointestinal tract of fish: biology, ontogeny, and environmental and nutritional modulation [J]. Reviews in Aquaculture, 2020, 12(2): 555-581. doi: 10.1111/raq.12340
[32] Zhu Y, Wang K, Jia X, et al. Antioxidant peptides, the guardian of life from oxidative stress [J]. Medicinal Research Reviews, 2024, 44(1): 275-364. doi: 10.1002/med.21986
[33] Chen Y, Liu E, Li C, et al. Effects of heat stress on histopathology, antioxidant enzymes, and transcriptomic profiles in gills of pikeperch Sander lucioperca [J]. Aquaculture, 2021, 534: 736277. doi: 10.1016/j.aquaculture.2020.736277
[34] Zou B, Xiao G, Xu Y, et al. Persimmon vinegar polyphenols protect against hydrogen peroxide-induced cellular oxidative stress via Nrf2signalling pathway [J]. Food Chemistry, 2018, 255: 23-30. doi: 10.1016/j.foodchem.2018.02.028
[35] Liu T, Li L, Yang Y, et al. Effects of chronic cold stress and thermal stress on growth performance, hepatic apoptosis, oxidative stress, immune response and gut microbiota of juvenile hybrid sturgeon (Acipenser baerii ♀× A. schrenkii♂) [J]. Fish & Shellfish Immunology, 2025, 157: 110078.
[36] Kampinga H H, Hageman J, Vos M J, et al. Guidelines for the nomenclature of the human heat shock proteins [J]. Cell Stress & Chaperones, 2009, 14(1): 105-111.
[37] Li L, Liu Z, Quan J, et al. Dietary nano-selenium alleviates heat stress-induced intestinal damage through affecting intestinal antioxidant capacity and microbiota in rainbow trout (Oncorhynchus mykiss) [J]. Fish & Shellfish Immunology, 2023, 133: 108537.
[38] Dawood M A O, Eweedah N M, Elbialy Z I, et al. Dietary sodium butyrate ameliorated the blood stress biomarkers, heat shock proteins, and immune response of Nile tilapia (Oreochromis niloticus) exposed to heat stress [J]. Journal of Thermal Biology, 2020, 88: 102500. doi: 10.1016/j.jtherbio.2019.102500
[39] Li S, Wang R, Dai Z, et al. Dietary supplementation with Yucca schidigera extract alleviated heat stress-induced unfolded protein response and oxidative stress in the intestine of Nile tilapia (Oreochromis niloticus) [J]. Ecotoxicology and Environmental Safety, 2021, 219: 112299. doi: 10.1016/j.ecoenv.2021.112299
[40] Yi D, Hou Y, Tan L, et al. N-acetylcysteine improves the growth performance and intestinal function in the heat-stressed broilers [J]. Animal Feed Science and Technology, 2016, 220: 83-92. doi: 10.1016/j.anifeedsci.2016.07.014
[41] Song Z H, Cheng K, Zheng X C, et al. Effects of dietary supplementation with enzymatically treated Artemisia annua on growth performance, intestinal morphology, digestive enzyme activities, immunity, and antioxidant capacity of heat-stressed broilers [J]. Poultry Science, 2018, 97(2): 430-437. doi: 10.3382/ps/pex312
[42] Rungrassamee W, Klanchui A, Maibunkaew S, et al. Bacterial dynamics in intestines of the black tiger shrimp and the Pacific white shrimp during Vibrio harveyi exposure [J]. Journal of Invertebrate Pathology, 2016, 133: 12-19. doi: 10.1016/j.jip.2015.11.004
[43] Cheng Y F, Chen Y P, Chen R, et al. Dietary mannan oligosaccharide ameliorates cyclic heat stress-induced damages on intestinal oxidative status and barrier integrity of broilers [J]. Poult Sci, 2019, 98(10): 4767-477. doi: 10.3382/ps/pez192


计量
- 文章访问数: 20
- HTML全文浏览量: 5
- PDF下载量: 2